

Motor-sensory convergence in tactile perception
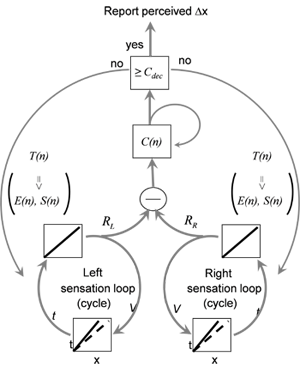
Two extreme possibilities are:
- motor and sensory signals interact within an open-loop scheme in which motor signals determine sensory sampling but are not affected by sensory processing.
- motor and sensory signals are affected by each other within a closed loop scheme.
We studied the scheme of motor-sensory interactions in humans using a novel object localization task that enabled monitoring the relevant overt motor and sensory variables. We found that motor variables were dynamically controlled within each perceptual trial, such that they gradually converged to steady values.
Training on this task resulted in improvement in perceptual acuity, which was achieved solely by changes in motor variables, without any change in the acuity of sensory readout. The within-trial dynamics is captured by a hierarchical closed loop model in which lower loops actively maintain constant sensory coding, and higher loops maintain constant sensory update flow.
These findings demonstrate interchangeability of motor and sensory variables in perception, motor convergence during perception and a consistent hierarchical closed loop perceptual model.
Active temporal coding
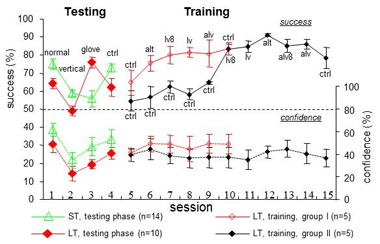
While scanning a textured surface with fingers, tactile information is encoded both spatially, by differential activation of adjacent receptors, and temporally, by changes in receptor activation during movements of the fingers across the surface.
We used a tactile discrimination task to examine the dependence of human tactile perception on the availability of spatial and temporal cues. Subjects discriminated between spatial frequencies of metal gratings presented simultaneously to both hands. Tactile temporal cues were eliminated by preventing lateral hand movements; tactile spatial cues were eliminated by using gloves with an attached rubber pin.
Analysis revealed separation of the subjects into two groups: “spatiotemporal” (ST) and ”latent-temporal” (LT). Under normal conditions, the performance of ST subjects was significantly better than that of the LT subjects. Prevention of lateral movements impaired performance of both ST and LT subjects. However, when only temporal cues were available, the performance of ST subjects was impaired, while that of the LT subjects either improved or did not change.
Under the latter condition, LT subjects changed strategy to scanning with alternating hands, at velocities similar to the velocities normally employed by ST subjects. These velocities generated temporal frequencies between 15 – 30 Hz. The LT subjects were unaware of their improved performance. Nine out of ten LT subjects significantly improved their performance under normal conditions when trained to scan gratings using alternating hands and velocities similar to those employed by ST subjects (Figure).
We conclude that:
- temporal cues are essential for spatial-frequency discrimination
- human subjects vary in the tactile strategies they use for texture exploration
- poor tactile performers can significantly improve by using strategies that emphasize temporal cues.
Relevant papers
-
 (2001). Temporal and spatial coding in the rat vibrissal system. Progress in brain research. $$1130.
(2001). Temporal and spatial coding in the rat vibrissal system. Progress in brain research. $$1130. -
Ahissar, E; Vaadia, E; Ahissar, M; Bergman, H; Arieli, A; Abeles, M (1992). Dependence of cortical plasticity on correlated activity of single neurons and on behavioral context. Science 1992 Sept 4; 257(5075):1412-5..
-
(1998). Temporal-code to rate-code conversion by neuronal phase-locked loops. Neural Computation. 10:597-650.
-
(2001). Figuring space by time. Neuron. 32:185-201. (Review)
-
(2001). Speech comprehension is correlated with temporal response patterns recorded from auditory cortex. Proceedings Of The National Academy Of Sciences Of The United States Of America. 98:13367-13372.
-
(2001). Temporal and spatial coding in the rat vibrissal system. Progress in brain research.
-
(2001). Importance of temporal cues for tactile spatial-frequency discrimination. Journal Of Neuroscience. 21:7416-7427.
-
(2006). Active sensation: insights from the rodent vibrissa sensorimotor system. Current Opinion In Neurobiology. 16:435-444.
-
(2006). Temporal decoding by phase-locked loops: Unique features of circuit-level implementations and their significance for vibrissal information processing. Neural Computation. 18:1611-1636.
-
(2008). Object localization with whiskers. Biological Cybernetics. 98:449-458. (Review)
-
(2008). 'Where' and 'what' in the whisker sensorimotor system. Nature Reviews Neuroscience. 9:601-612.
-
(2011). Motor-sensory convergence in object localization: a comparative study in rats and humans. Philosophical Transactions Of The Royal Society B-Biological Sciences. 366:3070-3076.
-
(2012). Seeing via miniature eye movements: a dynamic hypothesis for vision. Frontiers In Computational Neuroscience. 6.
-
(2012). Fast Feedback in Active Sensing: Touch-Induced Changes to Whisker-Object Interaction. Plos One. 7.
-
(2012). Motor-Sensory Confluence in Tactile Perception. Journal Of Neuroscience. 32:14022-14032.